


Surface structurale : 3. Description
De dimension modeste, l'ancienne carrière de Broussey-en-Blois présente un front de taille de quelques mètres de hauteur. Les conditions d'affleurement (présence de nombreux tas d'éboulis devant le front de taille et autour) sont comparables à celles de l'ancienne carrière de Saulvaux située à proximité.
Les roches, fortement diaclasées, sont disposées en strates horizontales de quelques décimètres d'épaisseur. Il s'agit d'une série homogène de calcaires micritiques de couleur beige à jaunâtre à patine noire ou parfois rouge (fig.2). Un niveau dolomitique laminé (roche de couleur rousse) est présent dans la partie supérieure du front de taille.
L'observation des éboulis n'a livré aucune trace de fossiles. L'occurence de quelques terriers trahit toutefois la présence de vie au moment du dépôt du sédiment à l'origine de la roche. Celle-ci résiste mal à l'action du gel (cf fiche Ménil-la-Horgne).

Fig.2 : Vue d'ensemble du front de taille de l'ancienne carrière de Broussey-en-Blois.
D'après la carte géologique, cette formation est à rattacher à l'Oxfordien supérieur (ex-Séquanien - fig.3).
Le faciès et la quasi-absence de faune caractérisent la formation des Calcaires à astartes. Le niveau dolomitique indique peut-être le sommet de la formation (cf. notice de la carte géologique de Commercy).
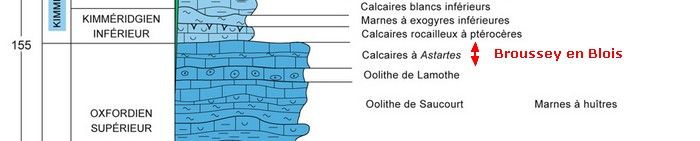
Fig.3 : Positionnement de la carrière dans la série stratigraphique (données chiffrées = âges en millions d'années - © BRGM).
Le plancher de la carrière est une surface structurale sur laquelle apparaissent des rides (ou ripple-marks), similaires à celles également observées dans la carrière de Saulvaux. Ces structures sédimentaires résultent de l'action de la houle dans un milieu peu profond (fig.4 et 5).

Fig.4 : Surface de base de la carrière décapée: ondulations symétriques = rides de houle

Fig.5 : Surface structurale et rides de houle en gros plan.
La présence de bioturbation (terriers de type Thalassinoides dus à l'action de crustacés - fig.6 et fiche Saulvaux) et de moules internes d'astartes (genre Nicaniella - cf. fiche Ménil-la-Horgne), bivalves endobiontes de petite taille à costulation concentrique ayant donné leur nom à la formation, constituent généralement les principales et rares traces de vie animale dans cette série sédimentaire étudiée en détail sur d'autres sites de la région (Lefort, 2012). Cette biodiversité, réduite à des associations fauniques monogénériques ou monospécifiques, traduit le plus souvent des conditions d'oxygénation limitée et/ou de salinité instable (Oschmann, 1994).
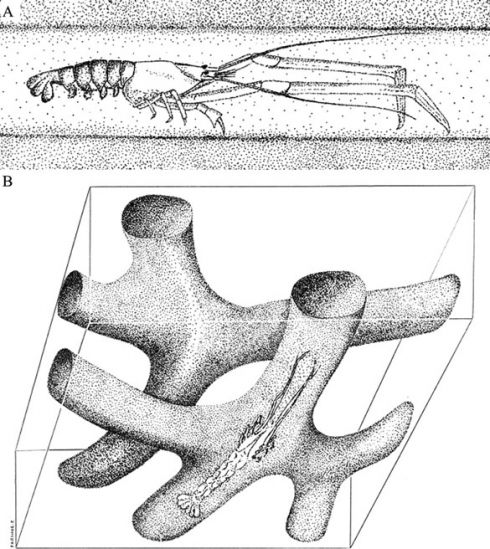
Fig.6 : Mecochirus rapax (A) un exemple de crustacé du Crétacé responsable du creusement de terriers de type Thalassinoides (B) comparables à ceux des Calcaires à astartes ( in Carvalho et al., 2007).
Les astartes, tout comme les crustacés responsables de la bioturbation, sont considérés comme des organismes se nourrissant par filtration du sédiment et tolérants vis-a-vis de l'oxygénation du milieu (Boyer et Droser, 2007; Oschmann, 1994; Savrda et Bottjer, 1989). Ils peuvent donc supporter un milieu pauvre en oxygène (= dysoxie).
Les astartes sont également sensibles aux variations de salinité. La petite taille de leur coquille serait à mettre en relation avec d'importantes variations de la salinité de l'eau (Fürsich et Werner, 1986).
La rareté des traces de vie dans les bancs de la carrière de Broussey est vraisemblablement le signe de conditions d'oxygénation (et de salinité ?) incompatibles avec la vie ou limitant fortement l'installation d'une biocénose diversifiée.
Faciès et contenu paléontologique des Calcaires à astartes permettent de caractériser un milieu de dépôt correspondant à un lagon très peu profond et bien abrité (Lefort, 2012).
Bibliographie
BOYER D. L. et DROSER M. L. (2007) - Devonian monospecific assemblages: new insights into the ecology of reduced-oxygen depositional settings. Lethaia, 40, pp. 321-333.
CARVALHO C.N., VIEGAS P.A. et CACHAO M. (2007) - Thalassinoides and its producer: Populations of Mecochirus buried within their burrow systems, Boca Do Chapim Formation (Lower Cretaceous), Portugal. Palaios, 22, pp.104-109.
FÜRSICH F. T. et WERNER W. (1986) - Benthic associations and their environmental significance in the Lusitanian Basin (Upper Jurassic, Portugal). Neues Jahrbuch für Geologie und Paläontologie Abhandlungen, Stuttgart, 172(3), pp. 271-329.
LEFORT A. (2012) - La limite Oxfordien-Kimméridgien: stratigraphie et paléoenvironnements dans les domaines téthysien (est et sud du Bassin de Paris, France) et boréal (Ile de Skye, Ecosse). Thèse de doctorat - Université Henri Poincaré, Nancy, 306 p.
OSCHMANN W. (1994) - Distribution, dynamics and palaeoecology of Kimmeridgian (Upper Jurassic) shelf anoxia in Western Europe. Modern and ancient continental shelf anoxia. Geological Society Special Publication, 58, pp. 381-395.
SAVRDA C. E. et BOTTJER D. J. (1989) - Trace-fossil model for reconstructing oxygenation histories of ancient marine bottom waters: application to Upper Cretaceous Niobrara Formation, Colorado. Palaeogeography, Palaeoclimatology, Palaeoecology, 74, pp. 49-74.
Auteurs : Philippe MARTIN - Didier ZANY - Date de création : 14/03/2013 - Dernière modification : 16/08/2013